- Mar 16, 2004
- 22,030
- 7,265
- 62
- Gender
- Male
- Faith
- Calvinist
- Marital Status
- Single
- Politics
- US-Democrat
I aked for what I should expect to find if I were looking for evidence of God in nature - you reply that evidence of God is "obvious" and that our failure to find it is due to our refusal to define "creation".
That's a bit of a non-answer. It's sounds as though you're saying "The evidence is so obvious that if fail to notice it's your own fault" - without specifying what this evidence actually is.
The Scriptures are evidence as I have made clear repeatedly, in accordance with the traditions of the faith and the teachings of the church for 2,000 years. If you are interested in an apologetic defense of the Scriptures I'm more then happy to provide one either here or in the formal debate area if you prefer.
God's is self existing and evidence for both the character and works of God remain self evident and obvious. With regards to revelation the testimony of Scripture stands on it's own merit, clearly indicating that our lineage and the lineage of all life on this planet begins with creation week, roughly 6,000 years ago.
IF it could be demonstrated that any complex organ existed, which could not possibly have been formed by numerous, successive, slight modifications, my theory would absolutely break down. (Charles Darwin, Origin of Species; VI. Difficulties of the Theory)
Here is such an organ, the size and complexity of the human brain.
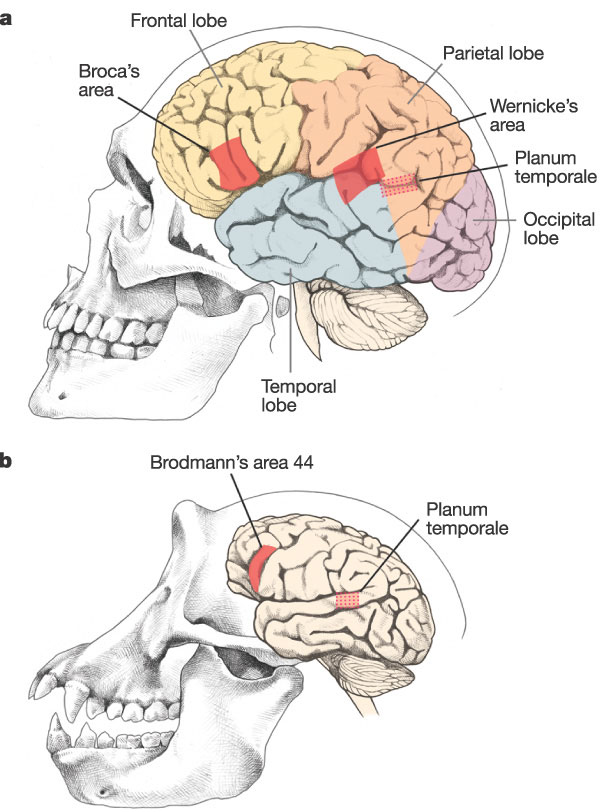
FIGURE 2. Comparative neuroanatomy of humans and chimpanzees. (Genetics and the making of Homo sapiens. Nature April 2003)
I think another user (Papias?) has answered your point on genetics and human EQ before; we already know of a few genes which increase human brain size and folding. A few that I know of include:
As for chimpanzee fossils, it's possible we will find them in the future. After all, we have found quite a few Miocene ape fossils - so not all primate fossils are automatically considered hominins.
- HAR1 (involved in the development of the cerebral cortex, and possibly sperm production)
- ASPM and CDK5RAP2 (both responsible for controling brain size).
Papias has never so much as commented on the HAR1f regulatory gene as many times as I have posted for several years now.
Here we report that the most dramatic of these ‘human accelerated regions’, HAR1, is part of a novel RNA gene (HAR1F) that is expressed specifically in Cajal– Retzius neurons in the developing human neocortex from 7 to 19 gestational weeks, a crucial period for cortical neuron specification and migration. HAR1F is co-expressed with reelin, a product of Cajal–Retzius neurons that is of fundamental importance in specifying the six-layer structure of the human cortex. HAR1 and the other human accelerated regions provide new candidates in the search for uniquely human biology."
The 118-bp HAR1 region showed the most dramatically accelerated change, with an estimated 18 substitutions in the human lineage since the human–chimpanzee ancestor, compared with the expected 0.27 substitutions on the basis of the slow rate of change in this region in other amniotes. Only two bases (out of 118) are changed between chimpanzee and chicken, indicating that the region was present and functional in our ancestor at least 310 million years (Myr) ago. No orthologue of HAR1 was detected in the frog (Xenopus tropicalis), any of the available fish genomes (zebrafish, Takifugu and Tetraodon), or in any invertebrate lineage, indicating that it originated no more than about 400Myr ago (Nature, 14 September 2006)
The 118-bp HAR1 region showed the most dramatically accelerated change, with an estimated 18 substitutions in the human lineage since the human–chimpanzee ancestor, compared with the expected 0.27 substitutions on the basis of the slow rate of change in this region in other amniotes. Only two bases (out of 118) are changed between chimpanzee and chicken, indicating that the region was present and functional in our ancestor at least 310 million years (Myr) ago. No orthologue of HAR1 was detected in the frog (Xenopus tropicalis), any of the available fish genomes (zebrafish, Takifugu and Tetraodon), or in any invertebrate lineage, indicating that it originated no more than about 400Myr ago (Nature, 14 September 2006)
What no researcher has been able to produce is an explanation for why and how a gene allowing only 2 substitutions in 400 million years suddenly gets 18, about 2 1/2 million years ago.
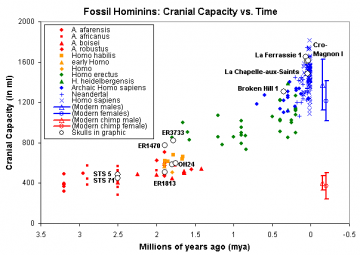
A. Afarensis with a cranial capacity of ~430cc lived about 3.5 mya.
A. Africanus with a cranial capacity of ~480cc lived 3.3-2.5 mya.
P. aethiopicus with a cranial capacity of 410cc lived about 2.5 mya.
P. boisei with a cranial capacity of 490-530cc lived between 2.3-1.2 mya.
OH 5 'Zinj" with a cranial capacity of 530cc lived 1.8 mya.
KNM ER 406 with a cranial capacity of 510cc lived 1.7 million years ago.
A. Africanus with a cranial capacity of ~480cc lived 3.3-2.5 mya.
P. aethiopicus with a cranial capacity of 410cc lived about 2.5 mya.
P. boisei with a cranial capacity of 490-530cc lived between 2.3-1.2 mya.
OH 5 'Zinj" with a cranial capacity of 530cc lived 1.8 mya.
KNM ER 406 with a cranial capacity of 510cc lived 1.7 million years ago.
Where are the ancestors for the Chimpanzees in all of this? No matter, there are many candidates in the list except that they are all regarded as our ancestors and none of them are considered Chimpanzee ancestors. The giant leap to cranial capacities approaching that of humans does not begin until just under 2 million years ago. I have long contended that there is neither the time nor the means for this transition.
"The skeleton was about 1.60 m (5 ft 3 in) tall, although he might have been 68 kg (150 lb) and 1.85 m (6 ft 1 in) tall had he lived to adulthood. The total skeleton is made up of 108 bones accounted for. The cranial capacity of Turkana Boy was about 880 cc, although if he had lived to adulthood, it would have been about 910 cc"
Turkana Boy, Wikipedia
Turkana Boy, Wikipedia
At adulthood Turkana Boy would have had a cranial capacity of 910cc or close to twice that of the African apes who are considered our ancestors. Then the cranial capacity remained static until 100,000 years ago. Let us not forget that by some other equally mysteries evolutionary process the Neanderthals developed a cranial capacity 10% greater then our own. That is in addition to the fact that we have no chimpanzee ancestors to compare the cranial capacity to. That is because every ape skull unearthed in Africa and Asia is automatically celebrated as one of our ancestors in a desperate attempt to find this mythical transitional apeman. Not once has Papias engaged me on this subject matter. Not once has he proven capable of doing the research and fielding the arguments he would have to.
Here is another list of hominid fossils regarded among creationists as human.
Hexian 412,000 years old had a cranial capacity of 1,025cc.
ZKD III (Skull E I) 423,000 years old had a cranial capacity of 915cc.
ZKD II (Skull D I) 585,000 years old had a cranial capacity of 1,020cc
ZKD X (Skull L I) 423,000 years ago had a cranial capacity of 1,225cc
ZKD XI (Skull L II) 423,000 years ago had a cranial capacity of 1,015cc
ZKD XII (Skull L III) 423,000 years ago had a cranial capacity of 1,030cc
Sm 3 >100,000 years ago had a cranial 917cc
KNM-WT 15000 (Turkana Boy) 1.5 million years ago had a cranial capacity of 880cc
(Source: Endocranial Cast of Hexian Homo erectus from South China, AMERICAN JOURNAL OF PHYSICAL ANTHROPOLOGY 130:445–454. 2006)
ZKD III (Skull E I) 423,000 years old had a cranial capacity of 915cc.
ZKD II (Skull D I) 585,000 years old had a cranial capacity of 1,020cc
ZKD X (Skull L I) 423,000 years ago had a cranial capacity of 1,225cc
ZKD XI (Skull L II) 423,000 years ago had a cranial capacity of 1,015cc
ZKD XII (Skull L III) 423,000 years ago had a cranial capacity of 1,030cc
Sm 3 >100,000 years ago had a cranial 917cc
KNM-WT 15000 (Turkana Boy) 1.5 million years ago had a cranial capacity of 880cc
(Source: Endocranial Cast of Hexian Homo erectus from South China, AMERICAN JOURNAL OF PHYSICAL ANTHROPOLOGY 130:445–454. 2006)
(My emphasis) Evolution is not atheisms' personal trump card, not matter how often they treat it as such. They deny theistic reasons because they are atheists, not because Darwinism is so damaging to religion.
I reject Darwinism because it lacks any substantive bearing on the genuine article of science. It's only purpose would appear to be to argue against creation. Never the less, Darwin himself proposed a logical disproof of his theory. I have an answer for that. In order to examine the scientific basis for common descent I propose to examine the genetic basis for the common descent of humans from that of apes. The most dramatic and crucial adaptation being the evolution of the human brain. With a cranial capacity nearly three times that of the chimpanzee the molecular basis for this giant leap in evolutionary history is still almost, completely unknown. Changes in brain related genes are characterized by debilitating disease and disorder and yet our decent from a common ancestor with the chimpanzee would have had to be marked by a massive overhaul of brain related genes. I propose that a critical examination of common descent in the light of modern insights into molecular mechanisms of inheritance is the single strongest argument against human/ape common ancestry.
Last edited:
Upvote
0

")

