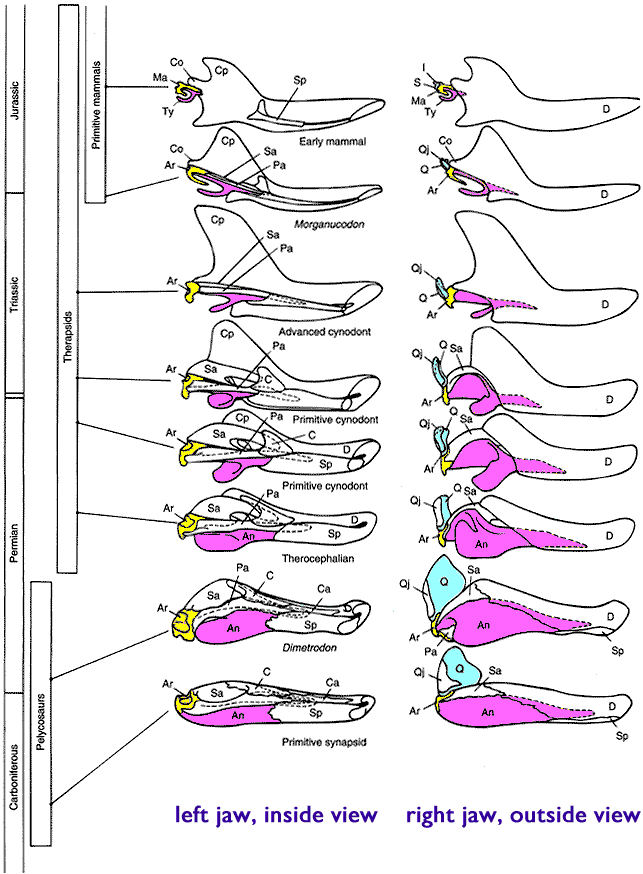
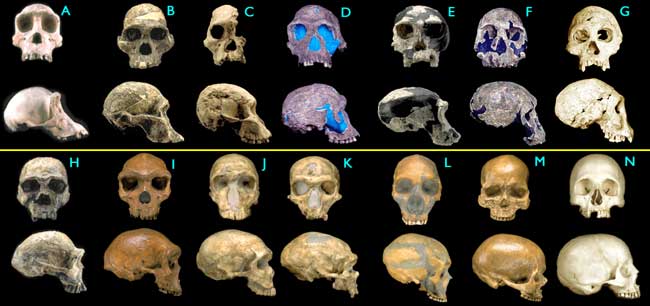
This is a topic from another forum, created entirely by someone that goes by the name of Reformentia. It is rather long, but conclusive. This will include many posts, so I would apreciate it if they were uninterupted, no posting untill I am dont please. From this sentence on, it is only his words.
This topic is not "evolution vs. creationism". This is "Here's the evidence for evolution, discuss". If you think it's incorrect or insufficient, feel free to share why. I am constantly seeing claims that there isn't any evidence for evolution occuring... like for example in the current "evolution vs. creationism" topic. This is a response to those claims.
This is a series of posts I made on the General forums of the game that shall not be named back about a year or so ago. It's my best shot at putting together a relatively comprehensive overview of the kind of evidence we're dealing with when we talk about evolution occuring and presenting it in a way which is hopefully fairly clear even to those who don't have much familiarity with the subject. Some visitors to the Legion offsites may have seen these there as well.
If anyone notices any broken links I'd appreciate having them pointed out. I think I've verified them all but I might have missed something.
Post 1: Dating Methods
Carbon (C14) Dating:
C14 dating is used to date the remains of organic, air breathing organisms up to approximately 50,000 years old. While living these organisms breathe the atmosphere, which contains trace amounts of the radioactive isotope Carbon 14 that is constantly being produced in the upper atmosphere through neutron bombardment. So long as they are alive the C14 content of their bodies will remain in equilibrium with the C14 content of the atmosphere. When they die respiration ceases, along with the intake of any new quantities of C14. Over time the C14 decays with a half-life of 5568 years into N14. By measuring how much C14 remains un-decayed the time elapsed since the death of the organism can be determined.
A common misperception of C14 dating is that it relies on the assumption that atmospheric C14 levels remained constant in the past so that we can know how much C14 an organism started off with. While this was an assumption made when the technique was first developed about half a century ago it has not been the case for several decades. Historical atmospheric C14 concentrations have been charted and calibrated using both dendochronology and lake varves which incorporate organic sediment in their annual deposition layers. One particularly good example of this is Lake Suigetsu in Japan where cores have been drilled to a depth of 45,000 annual layers. Because of the layering process we have an independent count of exactly how old every layer is and because the layers incorporate organic material (the remains of a surface algae which dies off every year and sinks to the bottom of the lake) each layer can be C14 dated as well, and using these two data points the atmospheric C14 content can be charted all the way back for the entire time span encompassed by the varve core. This data (cross-checked against multiple other sites and methods) then allows us to apply C14 dating to other sites already knowing how fluctuations in atmospheric C14 concentrations in the past will effect the results and allowing us to calibrate out error that would otherwise be introduced due to those past fluctuations.
Just one more note on C14 dating... once this calibration scale was applied it was discovered that previous C14 dates had been underestimating ages. By a few percent. There are also the occasional examples of C14 dates which have supposedly been wildly inaccurate. Many of these examples are the result of grossly improper applications of the method. For example, one I have encountered quite often is the "C14 dating of a living snail shell" that came back as thousands of years old... I believe this is one of Hovind's pet illustrations. The mollusks in question were extremely inappropriate subjects for C14 dating, which anyone familiar with the method would know. They form shells which are in equilibrium with the carbon content of the water sources in which they live... NOT the atmosphere. No C14 lab worth it's salt would ever date such an organism without warning the person requesting the test of the reservoir effect that would most likely render the test results invalid.
This topic is not "evolution vs. creationism". This is "Here's the evidence for evolution, discuss". If you think it's incorrect or insufficient, feel free to share why. I am constantly seeing claims that there isn't any evidence for evolution occuring... like for example in the current "evolution vs. creationism" topic. This is a response to those claims.
This is a series of posts I made on the General forums of the game that shall not be named back about a year or so ago. It's my best shot at putting together a relatively comprehensive overview of the kind of evidence we're dealing with when we talk about evolution occuring and presenting it in a way which is hopefully fairly clear even to those who don't have much familiarity with the subject. Some visitors to the Legion offsites may have seen these there as well.
If anyone notices any broken links I'd appreciate having them pointed out. I think I've verified them all but I might have missed something.
Post 1: Dating Methods
Carbon (C14) Dating:
C14 dating is used to date the remains of organic, air breathing organisms up to approximately 50,000 years old. While living these organisms breathe the atmosphere, which contains trace amounts of the radioactive isotope Carbon 14 that is constantly being produced in the upper atmosphere through neutron bombardment. So long as they are alive the C14 content of their bodies will remain in equilibrium with the C14 content of the atmosphere. When they die respiration ceases, along with the intake of any new quantities of C14. Over time the C14 decays with a half-life of 5568 years into N14. By measuring how much C14 remains un-decayed the time elapsed since the death of the organism can be determined.
A common misperception of C14 dating is that it relies on the assumption that atmospheric C14 levels remained constant in the past so that we can know how much C14 an organism started off with. While this was an assumption made when the technique was first developed about half a century ago it has not been the case for several decades. Historical atmospheric C14 concentrations have been charted and calibrated using both dendochronology and lake varves which incorporate organic sediment in their annual deposition layers. One particularly good example of this is Lake Suigetsu in Japan where cores have been drilled to a depth of 45,000 annual layers. Because of the layering process we have an independent count of exactly how old every layer is and because the layers incorporate organic material (the remains of a surface algae which dies off every year and sinks to the bottom of the lake) each layer can be C14 dated as well, and using these two data points the atmospheric C14 content can be charted all the way back for the entire time span encompassed by the varve core. This data (cross-checked against multiple other sites and methods) then allows us to apply C14 dating to other sites already knowing how fluctuations in atmospheric C14 concentrations in the past will effect the results and allowing us to calibrate out error that would otherwise be introduced due to those past fluctuations.
Just one more note on C14 dating... once this calibration scale was applied it was discovered that previous C14 dates had been underestimating ages. By a few percent. There are also the occasional examples of C14 dates which have supposedly been wildly inaccurate. Many of these examples are the result of grossly improper applications of the method. For example, one I have encountered quite often is the "C14 dating of a living snail shell" that came back as thousands of years old... I believe this is one of Hovind's pet illustrations. The mollusks in question were extremely inappropriate subjects for C14 dating, which anyone familiar with the method would know. They form shells which are in equilibrium with the carbon content of the water sources in which they live... NOT the atmosphere. No C14 lab worth it's salt would ever date such an organism without warning the person requesting the test of the reservoir effect that would most likely render the test results invalid.