Positive case, post 1.
Preliminary remarks
Thank you for your opening statement, Mark. I have a few observations.
I have included a “Terms” section to help us avoid confusion over exactly what certain terms mean.
The case for common ancestry from orthologous ERVs is not a mere homology argument. Read my post carefully, and you will see that it rests upon the probability of multiple separate coincidental integrations to corresponding DNA loci in different species being vanishingly small. I do not require your permission to disregard non-orthologous ERVs. They are essentially irrelevant.
I note that you cite certain items, but are inconsistent with citations and links. Links to tables and diagrams without an indication of the source documents are of little value, and you made an error with your link regarding CERV1 and PTERV1. You should have cited
(Polavarapu 2006) and not
(Bannert 2004). Please take a little more care with citations and links. On the subject of the suggestion that “at least one subfamily was present in the lineage prior to the time chimpanzees and humans diverged from a common ancestor”, three points: 1) This was based on estimated ages. 2) It is now recognized that the chimp/human split probably occurred over an extended period of time, and 3) these ERVs were not necessarily fixed (ubiquitous) in ancestral species. There are many other cases of genetic material “making it” into one lineage and not into a related lineage.
Terms
Exogenous retroviruses (XRVs) are infectious virus particles.
Endogenous retroviruses (ERVs) are reasoned to be exclusively the result of exogenous retroviral integrations with germ-line DNA (endogenizations). Exogenous retroviral integrations with somatic cell DNA are not endogenous. Their integrations are normally called
proviruses.
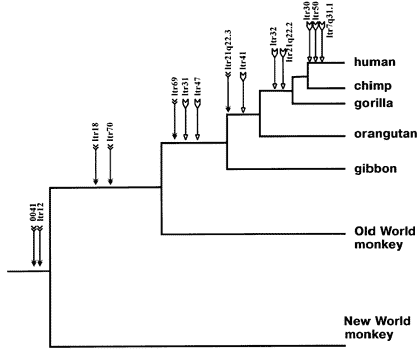
When we examine ERVs in different species, we may find them in corresponding locations in the DNA of those species. These are usually called
orthologous ERVs, orthologous meaning equivalent by way of being derived by common ancestry. As this is the question under discussion, I shall not use the word orthologous, so as not to beg the question. Instead, I will say “corresponding” or "shared”, and will then proceed to prove that corresponding ERVs are indeed orthologous.
Fixed ERVs are ERVs that are present in all individuals of a species.
Non-orthologous ERVs indicate separate endogenizations in different species, or the failure of an ERV to become fixed in one daughter species of a common ancestor, while becoming fixed, or at least represented in another daughter species. Non-corresponding (non-orthologous) ERVs are not evidence for common ancestry. Neither are they evidence for uncommon ancestry. They are essentially irrelevant to the case for common ancestry from corresponding ERVs. Perhaps an analogy will help here. Bill Bloggs is recorded on video, waving a gun around and stuffing dollar bills into his swag-bag at a branch of Wells Fargo. This is evidence that he is a bank robber. He is not recorded waving a gun around and stuffing dollar bills into his swag-bag at a branch of Citigroup, but this is
not evidence that he is
not a bank robber!.
Exogenous Retroviruses
Retroviruses enter host cells, “reverse” transcribe their RNA genomes into DNA, and integrate the resulting DNA with the host cell’s nuclear DNA.
(Temin, 1970),
(Baltimore, 1970),
(Coffin, 1979).
Although there are general statistical trends the locus of the integration is highly variable. Retroviruses tend to target particular cell types, but do not infect all of them.
(Taruscio 1991),
(Skinner et al., 2001),
(Mitchell, 2004),
(Wang et al., 2007).
The Wang study showed that the probability of a retrovirus endogenizing in the same location twice is no greater than 1/500.
Endogenous Retroviruses.
The structure of a provirus and a complete* ERV are essentially the same. Most ERVs, however, are not fully functional proviruses.
(Coffin, 1979),
(Weiss, 2006)
* There are many fragments of ERVs in our DNA, and in that of chimpanzees.
An example of a retrovirus in the process of becoming endogenized is the koala virus KoRV.
(Tarlinton, 2006). A functional retrovirus, the “Phoenix Virus” has been resurrected from dormant ERVs.
(Dewannieux 2006). These studies confirm the retroviral origins of ERVs.
In contrast with provirus integrations in somatic cells, ERVs are found in the DNA of all nuclear cells, Each ERV being found in exactly the same location, going from cell to cell. This is why they are regarded as being endogenous. All the cells of the body are daughter cells of the zygote, formed by the fusion of the sex gametes. Going back through our ancestry, for each type of ERV, there has to have been a single ancestral gamete or zygote that an exogenous retrovirus endogenized.
(International Human Genome Sequencing Consortium, 2001). ERVs may multiply further in the DNA by retrotransposition, but these may also be regarded as additional instances of endogenization - they have to have occurred in germline cells in order to be endogenous.
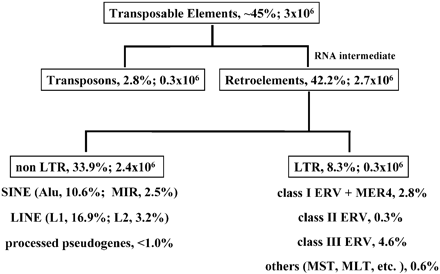
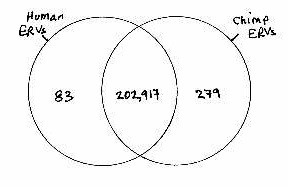
There are some 200,000 ERVs and ERV fragments in the human and the chimpanzee genomes. Less than 1% of these are lineage-specific (no corresponding ERV in the other species).
(International Human Genome Sequencing Consortium, 2001),
(Chimpanzee Sequencing and Analysis Consortium, 2005).
For all these ERVs and ERV fragments to have endogenized separately in each species, but for their locations to correspond as they do, would be a coincidence of truly staggering unlikelihood. For one pair of endogenizations to correspond by coincidence, the probability would be 1/500. For two, it would be 1/500 x 1/500 = 1/250,000. For three, 1/500 x 1/500 x 1/500 = 1/12,500,000 or 1/500[SUP]3[/SUP]. For 200,000, the probability of coincidence is 1/500[SUP]200,000[/SUP]. For all intents and purposes, it is impossible for chimps and humans
not to have had common ancestors.
References
Baltimore, D. “Viral RNA-dependent DNA Polymerase” Nature vol 226, p1209 June 27 1970.
http://tamu.edu/faculty/kunkel/Baltimore.pdf
The Chimpanzee Sequencing and Analysis Consortium, “Initial sequence of the chimpanzee genome and comparison with the human genome“, Nature 437, 69-87 (1 September 2005),
Initial sequence of the chimpanzee genome and comparison with the human genome : Article : Nature
Coffin, J. M. “Structure, Replication, and Recombination of Retrovirus Genomes: Some Unifying Hypotheses”, J. gen. Virol 1979, 42, 1-26.
http://jgv.sgmjournals.org/content/42/1/1.full.pdf
Dewannieux, M., Harper, F., Richaud, A., Letzelter, C., Ribet, D., Pierron, G., Heidmann, T., “Identification of an infectious progenitor for the multiple-copy HERV-K human endogenous retroelements”, Genome Res. 2006, 16: gr.5565706.
Identification of an infectious progenitor for the multiple-copy HERV-K human endogenous retroelements
International Human Genome Sequencing Consortium, “Initial sequencing and analysis of the human genome”, Nature 409, 860-921 (15 February 2001).
Initial sequencing and analysis of the human genome : Article : Nature
Mitchell, R.S., Beitzel, B.F., Bushman, F.D., “Retroviral DNA Integration: ASLV, HIV, and MLV Show Distinct Target Site Preferences”, PLoS Biol. 2004 August 2(8): e234.
Retroviral DNA Integration: ASLV, HIV, and MLV Show Distinct Target Site Preferences
Skinner, L. M., Sudol, M., Harper, A. L., Katzman, M., “Nucleophile Selection for the Endonuclease Activities of Human, Ovine, and Avian Retroviral Integrases” The Journal of Biological Chemistry, January 5, 2001 276, 114-124.
Nucleophile Selection for the Endonuclease Activities of Human, Ovine, and Avian Retroviral Integrases
Tarlinton R.E., Meers, K., Young, P. R. “Retroviral invasion of the koala genome“ Nature 442, 79-81 (6 July 2006).
http://www.nature.com/nature/journal/v442/n7098/full/nature04841.html
Taruscio, D., Manuelidis, L., “Integration site preferences of endogenous retroviruses.“ PubMed Chromosoma. 1991 Dec;101(3):141-56.
Integration site preferences of endogenous retrov... [Chromosoma. 1991] - PubMed - NCBI
Temin H.M. and Mizutani, S. “RNA-Dependent DNA Polymerase in Virions of Rous Sarcoma Virus”, Nature vol. 226 p1211 June 27 1970.
http://tamu.edu/faculty/kunkel/Temin.pdf
Wang, G. P., Ciuffi, A., Bushman, F. D., “HIV integration site selection: Analysis by massively parallel pyrosequencing reveals association with epigenetic modifications”, Genome Res. 2007 August; 17(8): 1186-1194.
HIV integration site selection: Analysis by massively parallel pyrosequencing reveals association with epigenetic modifications
Weiss, R. A., “The Discovery of Endogenous Retroviruses”, Retrovirologt 2006 3:67.
http://link.springer.com/content/pdf/10.1186/1742-4690-3-67.pdf